1. UVODglej predavanje Rast in razvoj - uvod, 2.del
Zmogljivost rasti ni med lastnostmi, ki bi ji pri selekciji prašičev odmerjali pozornost. To je tudi razumljivo, ker je zaradi velike nagnjenosti prašičev k zamastitvi, gospodarnost določala take klavne mase za katere ni potrebna večja zmogljivost rasti. V zadnjih dveh-treh desetletjih je uspelo s selekcijo prašiče dodobra razmastiti. Nekateri so pri tem že mnenja, da je doseglo maščevje svojo kritično mejo, ki jo zaradi slabšanja odpornosti in vitalnosti, ne kaže prestopiti. V tem je tudi eden od razlogov zakaj iščemo nove možnosti za izboljšanje gospodarnosti prireje prašičev tudi v večji masi ob zakolu. Prednosti težjega klavnega prašiča so v težji in primernejši šunki (pršut!), V bolj doraslih klavnih delih in v zmanjšanju stroškov za prirejo pujskov - na slovenskih farmah bi za isto prirejo žive mase ob povečanju klavne mase od 100 na 110 kg potrebovali 1750 svinj manj! Veliki pomankljivosti pa sta poslabšanje sestave telesa in povečanje porabe krme za enoto prirasta. Za pitanje na večjo maso so primerni prašiči z veliko zmogljivostjo rasti, s specifičnim potekom rasti in z izraženo sposobnostjo nadomestne rasti. Rast prašičev bi morala biti zmerna pri nizkih masah in hitra proti koncu pitanja ne da bi pri tem prišlo do prevelike zamastitve. Pri takem poteku rasti je z vso upravičenostjo mogoče pričakovati tudi trdnejše in močnejše noge, ker bi imelo kostno tkivo daljše obdobje svoje maksimalne rasti.
V Sloveniji razmišljanja o večji klavni masi niso od danes. V drugi polovici sedemdesetih let smo opravili v Sloveniji obsežne raziskave rasti prašičev in med drugim ugotovili pasemsko značilne poteke rasti ter značilne razlike v nadomestni rasti. Rezultate smo uporabili tudi pri selekciji merjascev iz testiranja, ko smo kriterije odbire pri 60 kg usklajevali po medpasemskih razlikah v poteku rasti. Prvi predlog za podaljšanje obdobja testiranja smo izdelali v letu 1985. Po obsežnih raziskavah rasti prašičev do mase 120 kg v letu 1988 smo obdobje testiranja prašičev za večino terminalnih pasem s 1.1.1989. Podaljšali na 30 - 120 kg. Ob tem za slovensko prašičerejo pomembnem ukrepu smatramo za koristno in potrebno, da pojasnimo nekatere biološke in gospodarske osnove selekcije prašičev na večjo zmogljivost rasti.
2. OSNOVNA BIOLOŠKA IN FIZIOLOŠKA SPOZNANJA O POSTNATALNI RASTI PRAŠIČEV.
Pred skoraj petdesetimi leti je McMEEKAN (1940,1941) s svojo znamenito raziskavo postnatalne rasti prašičev in objavo rezultatov v delu "GROWTH AND DEVELOPMENT IN THE PIG, WITH SPECIAL REFERENCE TO CARCASS QUALITY CHARACTERS" postavil temeljna spoznanja o biologiji postnatalne rasti prašiča. Hammondov model delitve hranil iz leta 1952 in Palssonov model diferencialne rasti iz leta 1955 sta njegova spoznanja potrdila in dopolnila.
HAMMOND (1952)-cit.po STEELE-u in sod.(1987)- je pojasnil različno rast posameznih telesnih tkiv kot posledico prioritete v prehrani ali kakor je drugače rečeno v "nutrition partitioning" (delitvi hranil). Njegov model pojasnuje, katero tkivo je bolj kritično za preživetje (živčevje > kosti > mišičje > maščobno tkivo) in da je prioriteta v prehrani pozitivno povezana z metabolično aktivnostjo tkiva. (shema 1)
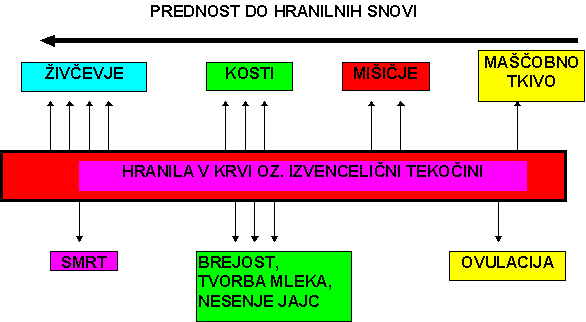
PALSSON (1955) je shematično (shema 2) predstavil vrstni red maksimalne rasti posameznih telesnih tkiv, ki ga ni mogoče zamenjati. Spreminjamo lahko le trajanje obdobja maksimalne rasti. Po tem vrstnem redu je maščobno tkivo zadnje na vrsti in ima tako najmanjšo prednost v prehrani. Posledica te diferencialne rasti telesnih tkiv je različna sestava dnevnega prirasta in s tem stalna spreminjanja sestave telesa. Shematsko je to pokazano v shemi 3, ki je napravljena po podatkih raziskave, ki so jo opravili METZ in dr. (1980).
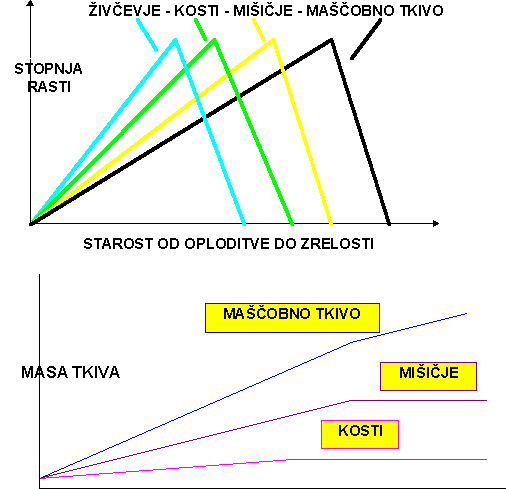
V shemah 2 in 3 je potrjen vrstni red obdobij maksimalne rasti posameznih telesnih tkiv. Rast posameznega telesnega tkiva je povezana z rastjo drugega tkiva.Kakor je poznano iz fiziologije rasti vodijo rast posameznih telesnih tkiv hormoni. O delovanju hormonov je še marsikaj nepojasnenega. V zadnjem času, ko so v ospredju raziskave s somatotropnim hormonom pa je praktično dnevno kaj novega.V tabeli 1 je, povzeto po KARG-u (1987), pokazano multifaktorielno delovanje hormonov na rast.
|
|
|
|
|
| insulin | pankreas | +* | + |
| glukagon | pankreas | (-) | (-) |
| GRH | hipotalamus | (+) | (-) |
| somatotropin | hipofiza | + | - |
| IGF n | jetra | + | (+) |
| somatostatin | hipotalamus, pankreas | - | (-) |
| adrenalin | adrenalka | (+) | - |
| noradrenalin | adrenalka | (+) | - |
| tiroksin | ščitnica : | . | . |
| . | nizke doze | + | (-) |
| . | visoke doze | - | - |
| glukokortikoidi | adrenalka : | . | . |
| . | nizke doze | (+) | (+) |
| . | visoke doze | - | - |
| androgeni | gonade | + | (-) |
| estrogeni | gonade, placenta : | . | . |
| . | nizke doze | + | (-) |
| . | visoke doze | - | + |
Delovanje somatotropnega hormona (somatotropina) je med drugimi shematično upodobil KARG (1987) in pokazano je v shemi 4.* + = ANABOLNI VPLIV
- = KATABOLNI VPLIV
( )= VERJETEN VPLIV
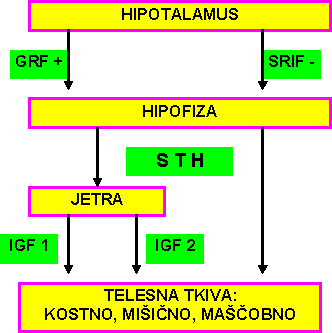
Somatotropin (STH) se izloča v hipofizi pod stimulativnim vplivom somatokrinina (GRH) in zavirujočim delovanjem somatostatina (SRIH). STH deluje na izločanje somatomedinov (IGFn) v jetrih, ki skupaj z insulinom delujejo na rast tkiv in skupaj s hormoni ščitnice na rast skeleta. STH neposredno vpliva na lipolizo in mobilizacijo glukoze.O ugodnem delovanju ekstraktov hipofize na rast podgan poročajo že EVANS in sod.(1931).V zadnjih letih so odkriti novi biotehnološki postopki pridobivanja somatotropina, kar je spodbudilo tudi številne raziskave vpliva STH na rast in sestavo telesa prašičev.Med njimi smo izbrali eno zadnjih, ki so jo objavili EVOCK in sod.(1988) in je pokazana v tabeli 2.
| lastnost | doza ppSTH* | . | . |
|
|
|
|
|
| končna masa (kg) |
|
|
|
| dnevni prirast (kg) |
|
|
|
| konverzija krme (kg) |
|
|
|
| dnevna konzumacija krme (kg) |
|
|
|
| randman (%) |
|
|
|
| debelina hrbtne slanine (cm) |
|
|
|
| masa šunke - meso s kostmi (kg) |
|
|
|
| površina hrbtne mišice (cm2) |
|
|
|
| mišičje (kg) |
|
|
|
| adipozno tkivo (kg) |
|
|
|
| jetra (kg) |
|
|
|
| ledvice (g) |
|
|
|
| osteohondroza (%) |
|
|
|
Podatki v tabeli 2 nedvoumno potrjujejo neposreden vpliv ppSTH na rast in sestavo telesa. Mimogrede povedano, ti rezultati tudi nakazujejo zelo verjeten razvoj bodočih postopkov prireje prašičjega mesa. Pri iskanju razlage gornjih sprememb ni mogoče mimo celične osnove postnatalne rasti. Zmanjšanje zamaščenosti je lahko posledica zmanjšanja števila adipocit oziroma njihove velikosti. V kolikor velja predvsem druga možnost, to pomeni, da s selekcijo na večjo mesnatost izbiramo živali pri katerih nastopi obdobje maksimalne rasti adipoznega tkiva kasneje. Posredno potrjujejo to podatki raziskave, ki so jo opravili SOLOMON IN SOD.(1988). Ugotovili so, da dodajanje pSTH vpliva na hipertrofijo mišičnih vlaken.* MIKROGRAMI ppSTH / KG TELESNE MASE / DAN
3. SELEKCIJA POTEKA IN ZMOGLJIVOSTI RASTI PRAŠIČEV
V literaturi je le nekaj informacij o selekciji poteka in zmogljivosti rasti živali. Eisen (1976) poroča, da so sicer maloštevilni poskusi pri miših pokazali srednjo uspešnost selekcije spremembe poteka rasti krivulje. Šalehar (1978, 1979) je v raziskavah rasti prašičev potrdil razlike v poteku rasti med pasmami in opredelil njegov pomen za selekcijo rasti prašičev. Krieter (1987) je na vsakoletnem simpoziju kmetijske fakultete v Kielu v predavanju "Zuchterische Moglichkeiten zur Regulation des Wachtstums beim Schwein" predstavil zanimive predhodne rezultate raziskave rasti prašičev nemška plemenita in Pietrain. Srednje vrednosti za parametre rastne krivulje in nekatere mere rasti pri obeh pasmah so v tabeli 3.
| . |
|
|
| končna masa (kg) |
|
|
| merilo zrelosti |
|
|
| masa pri max prirastu (kg) |
|
|
| starost pri max prirastu (teden) |
|
|
| max prirast pri infleksiji (kg/teden) |
|
|
Končna masa je med pasmami različna. Pasma pietrain je primer rano zrele in bela plemenita pozno zrele pasme. To potrjujejo tudi podatki za mere rasti in mere rastne krivulje.
Tabela 4: Korelacije med parametri krivulje in dnevnim prirastom (Krieter, 1988)
A k y t v Dnevni prirast
A - -.82 .82 .76 -.02 -.20
k - -.47 -.57 .52 .68
y - .91 .36 .02
t - .08 -.19
v - .83
A - končna masa
k - merilo zrelosti
y - masa pri maks. prirastu
t - starost pri maks. prirastu
v - maks. prirast pri inflekcijiMed dnevnim prirastom in končno maso ter starostjo pri maksimalnem prirastu je negativna korelacija. Živali z veliko zmogljivostjo rasti - veliko končno maso, so težje in starejše pri maksimalnem prirastu. Visoki in pozitivni korelaciji sta med dnevnim prirastom in merilom zrelosti ter maksimalnem prirastu pri inflekciji. To omogoča sklepanje na splošno manjšo rastnost živali z veliko zmogljivostjo rasti.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Končna masa in masa pri maksimalnem prirastu sta s masami pri 100 in 150 dneh v negativni ter s maso pri 200 dneh v pozitivni korelaciji. Živali z manjšo končno maso so težje pri starosti 100 oziroma 150 dni. Rano zrele živali imajo večje vrednosti za merilo zrelosti, kar potrjujejo tudi pozitivne korelacije s masami pri 100, 150 oziroma 200 dni. Negativne korelacije starosti pri maksimalnem prirastu, oziroma pozitivne maksimalnega prirasta pri inflekciji z masami pri 100, 150 oziroma 200 dneh ter značilno zmanjševanje v prvem oziroma povečevanje v drugem primeru samo še dodatno potrjuje, da je potek rasti rano oziroma poznozrelih pasem povsem različen.Krieter je pomen ugotovljenih povezav za praktično delo pri selekciji prašičev opredelil z naslednjim:
a. Večja končna masa ima pri svinjah za posledico zakasnitev spolne zrelosti. Zaradi restriktivne prehrane ter relativno kratke dobe rabe svinj pa to za rejo svinj nima večjega pomena.
b. Naslednje vprašanje je povezano s pomenom spremembe mase pri maksimalnem prirastu, če ostane končna masa pri pitanju nespremenjena. Vprašanje je upodobil s spodnjimi krivuljami, pri čemer krivulja št. 2 predstavlja sedanji potek dnevnega prirasta.
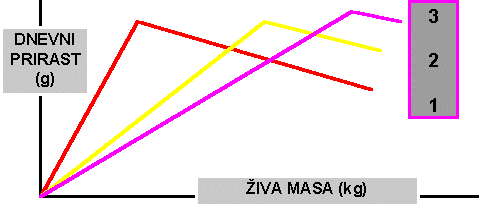
Z zmanjšanjem dnevnih prirastov po dosegu inflekcije nastopi močna zamasitev (krivulja št. 1). Obratno to pomeni, da je s premaknitvijo maksimalnega prirasta na višjo starost oziroma maso mogoče omejiti rast maščobnega tkiva.c) Selekcija na "zrelost" je mogoča:
- z neposredno odbiro na maso oziroma starost pri maksimalnem prirastu (praktično težko izvedljiva),
- s spremembo velikosti telesa (zaradi manjše povezave med mesnatostjo, rastnostjo, sestavo telesa in telesnimi merami ta pot ni mnogo obetajoča),
- s spremembo mase ali starosti živali pri odbiri - selekcija na živali, ki imajo rastno krivuljo tipa št. 3 (taka odbira utegne spremeniti začetno rast).
Zanimiva razmišljanja in rezultate je potrebno obravnavati kot predhodne in kakor pravi Krieter sam, tudi potrditi v obsežnejših poskusih.
4. GOSPODARNOST PITANJA NA VEČJO MASO
Gunter in sod. (1987) so upodobili povezavo med končno maso pri pitanju in potrebnim številom svinj (graf. 2).
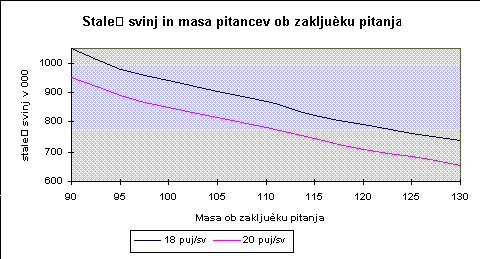
Zmanjšanje stroškov za prirejo pujskov pri večjih končnih masah pitancev pa sodeč po drugem delu Gunter-ja in sod. (1988) ne pokrije večjih stroškov pitanja. Svetujejo zmanjšanje končne mase pitancev - od sedanjih 120 kg na 105 do 110 kg.Zanimiva je raziskava, ki so jo na to temo izvedli Wiesemueller in sod. (1981). Ugotovili so, da je pri sedanjih prašičih intenzivna rast mišičja tudi še pri masah 115 do 120 kg. Poraba energije za enoto prirasta je pri pitanju na 120 kg oziroma 110 kg praktično enaka. Za zmanjšanje obsega investicij in za preprečitev prevelike gostote naselitve pitancev priporočajo pitanje do mase 115 do 120 kg. Ob tem še informirajo, da so v teku prehranski poskusi za optimiranje poteka rasti. Kovacs in dr. (1987) vidijo v povečanju končne mase možnost, da lahko s petimi pitanci s maso 120 kg priredijo enako količino mesa s kostmi kot s šestimi 100 kg prašiči.
Van Aelten in Sterkens (1984) sta proučila vpliv končne mase pitancev na gospodarnost pitanja, ki sta jo izrazila s pomočjo produkcijskega indeksa (PI):
PI* = KT x KV - PK x CK - TP x FS - (ZT - 20) x 2,7 x CK
KT- klavna masa
KV - klavna vrednost
PK - poraba krme
CK - vrednost 1 kg krme
TP - trajanje pitanja
FS - fiksni stroški na dan
ZT - začetna masa*Stroški za pujska niso vključeni.
V poskusu sta pitala prašiče do klavnih tež od 90 do 115 kg in izmerila pitovne ter klavne lastnosti (tab. 6).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tabela 7: Gospodarnost pri pitanju do različne končne mase (Van Aelten in dr. 1984)Podobno kot so v tovrstnih raziskavah ugotovili drugi, sta tudi van Aelten in Sterkens potrdila poslabšanje konverzije krme, zmanjšanje mesnatosti ob povečanju debeline hrbtne slanine in poslabšanje razmerja meso:slanina pri pitanju na večje mase. Na osnovi teh podatkov sta izračunala po svojem postopku gospodarnost, ki je pokazana v tabeli 7.* Belgijski franki
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rezultati kažejo, da se dohodek na prašiča oziroma stojišče povečuje s maso ob zakolu. Pri tem ni upoštevan strošek za pujska, kar bi ugoden izračun za večje mase ob zakolu samo še izboljšalo.5. ZAKLJUČEK
Zmogljivost rasti prašičev je ena izmed premalo izkoriščenih možnosti za gospodarnejšo prirejo prašičev. Biologija postnatalne rasti prašičev kaže na možnosti uravnavanja trajanja obdobij maksimalne rasti posameznega telesnega tkiva. Prašiči z večjo zmogljivostjo rasti imajo obdobje največje rasti pri višjih starostih oz. masah ter značilen potek rasti. S testiranjem prašičev do večjih mas lahko poiščemo živali, ki imajo veliko splošno zmogljivost rasti.Pri tem pa morajo imeti tudi veliko zmogljivost rasti mišičja in kasen nastop obdobja maksimalne rasti maščobnega tkiva. Zaradi verjetne povezave med zmogljivostjo rasti in plodnostjo ter potrebne specializacije pasem, ki prinaša večjo učinkovitost selekcije, ne kaže vseh pasem testirati do iste končne mase. Gospodarnost reje prašičev je potrebno iskati tudi v optimalni masi ob zakolu, ki mora biti vsklajena z zmogljivostjo rasti in s sposobnostjo nadomestne rasti.
6. LITERATURA
Literatura je na voljo pri avtorju.