|
|
|
|
|
|
|
|
. | . | . | . |
|
|
|
|
|
|
|
|
|
|
|
|
V - VISOKO ENERGETSKA KRMA
N - NIZKO ENERGETSKA KRMA
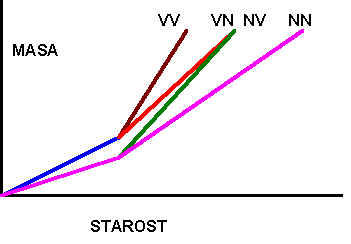
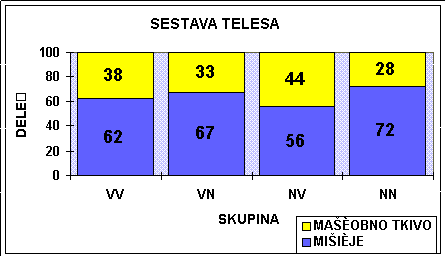
MODEL DELITVE HRANIL (HAMMOND 1952) Tabela : Multifaktorielno delovanje hormonov na rast (Karg, 1987)
Shema : Delovanje somatotropina, Karg 1987
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
POMEN:SOMATOTROPIN (STH) DELOVANJE STH:L. 1908 - Hipofiza in rast L. 1922 - Prvi poskusi z ekstrakti hipofize na podganah - povečanje rasti in spremembe sestave telesa (prerazporeditev hranil) UPORABA:Tvorba somatomedinov v jetrih Lipoliza, mobilizacija glukoze in ???? Biotehnološki postopki pridibivanja STH Subkutana aplikacija (dnevna aplikacija - boljša rast, konverzija krme, večja mesnatost, .......) BODOČA TEHNOLOGIJA ???????
Somatotropin :Najpomembnejši hormon rasti Učinki: somatogeni in metabolni SEKRECIJA SOMATOTROPINA
|
|
|
čas |
čas |
Mehanizmi delovanja STH :metabolni in somatogeni vplivi neposredni in posredni vplivi SOMATOGENI VPLIVI STH TEORIJA DVOJNEGA VPLIVA:
NEPOSREDNI VPLIV (dokaz: prisotnost celičnih receptorjev)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Lastnost/doza | 0* | 35* | 70* |
| Končna masa (kg) | 97 | 112 | 109 |
| Dnevni prirast (g) | 880 | 1010 | 980 |
| Konverzija krme (kg) | 3.3 | 2.8 | 2.5 |
| Dnevna konzumacija krme (kg) | 3.0 | 2.9 | 2.5 |
| Randma (%) | 72.7 | 72.1 | 70.7 |
| Debelina hrbtne slanine (cm) | 2.4 | 2.2 | 1.0 |
| Teža šunke . meso s kostmi (kg) | 7.6 | 9.5 | 9.3 |
| Površina hrbtne mišice (cm2) | 28 | 34 | 36 |
| Mišičje (kg) | 34.1 | 42.5 | 42.9 |
| Maščobno tkivo (kg) | 16.9 | 15.1 | 8.7 |
| Jetra (kg) | 1,8 | 2.1 | 2.3 |
| Ledvice (g) | 169 | 217 | 286 |
| Osteochondroza (%) | 1/8 | 4/8 | 4/8 |
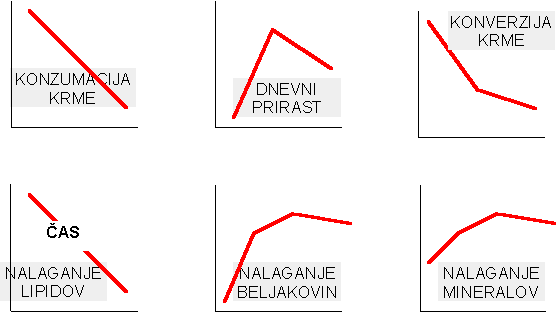
| Lastnost | Kontrola | pGH (100 ľg/kg/dan) |
| Nalaganje beljakovin (g/dan) | . | . |
| Merjasci | 196 | 238 |
| Svinjke | 148 | 235 |
| Kastrati | 139 | 225 |
| Nalaganje beljakovin (% od kontole) | . | . |
| Merjasci | 100 | 121 |
| Svinjke | 100 | 159 |
| Kastrati | 100 | 162 |
| Nalaganje maščob (g/dan) | . | . |
| Merjasci | 317 | 203 |
| Svinjke | 411 | 185 |
| Kastrati | 462 | 223 |
| Nalaganje maščob (% od kontrole) | . | . |
| Merjasci | 100 | 64 |
| Svinjke | 100 | 46 |
| Kastrati | 100 | 48 |
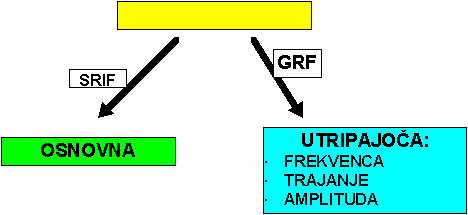
IZVOR: hipotalamusSOMATOSTATIN (SRIH)
DELOVANJE SRIH: redukcija nivoja STH, insulina in glukagona; supresivno delovanje na absorbcijo hranil v črevesju
NEVTRALIZACIJA DELOVANJA SRIH: aktivna imunizacija - povečanje rasti za 15 - 20% (raziskovalni nivo)SOMATOMEDINI - IGF1, IGF2 IZVOR: jetra pod vplivom STH, druga tkiva
POMEMBNEJŠA DELOVANJA: stimulira proliferacijo celic, vpliva na sprejem beljakovin in glukoze v mišicah in drugih tkivih; inhibira lipolizo
IGF1: vpliva postnatalno rast
IGF2: vpliva fetalno in neonatalno rast - dolga razpolovna doba, manjša nihanja v koncentraciji v plazmi.RASTNI FAKTORJI INSULINU PODOBEN RASTNI FAKTOR 1, 2 (IGF1, IGF2) NERVNI RASTNI FAKTOR (NGF): stimulira rast simpatičnih in senzoričnih živcev v embrijih, postanatalno simpatično živčevje EPIDERMALNI RASTNI FAKTOR (EGF): stimulira rast epitelnih celic, verjetno sodeluje pri regeneraciji živčevja, celjenju kit, angiogenezi
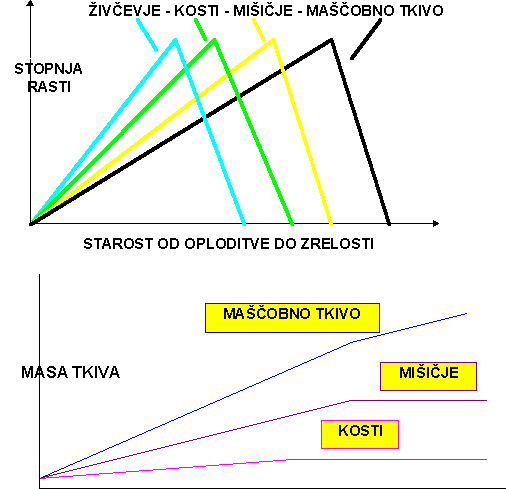
Posamezna tkiva različno rastejo v posameznih obdobjih rasti. Najprej raste skelet, nato mišičje in nazadnje maščobno tkivo. Obdobja rasti posameznih tkiv se v precejšnji meri prekrivajo in je to pravzaprav bolj vprašanje relativnih časov razvoja. V mladosti najprej najbolj intenzivno raste skelet, čeprav v tem času raste tudi mišičevje in se naloži nekaj maščob. Ko skelet zaključi z rastjo, mišičevje raste naprej. Ko to doseže svoj polno razvoj, se začne nalagati pravzaprav skoraj čista maščoba. Ta proces je mogoče skrajšati ali podaljšati s prehrano in selekcijo živali, ni pa mogoče zamenjati oziroma spremeniti vrstnega reda dogajanja (rasti posameznih tkiv).
Razmerje v velikosti telesnih organov ostanejo le redkokdaj enaka vse prenatalno in postnatalno življenje. Različni centri rasti telesa postanejo aktivni v različnem času in so različno aktivni, so koordinirani in so značilnost vrst (rečemo: maksimalna rast posameznih telesnih delov, organov ali tkiv se prične po natančno določenem vrstnem redu). Vsi deli živali ne prenehajo rasti hkrati; nekatera tkiva rastejo celo življenje. Telesni organi in tkiva rastejo z različno karakteristično stopnjo do končne velikosti; nekateri rastejo prej v življenju, medtem ko prično drugi rasti pozneje in pozno dosežejo svojo maksimalno velikost. Stopnja rasti vsakega organa oziroma tkiva narašča do maksimuma, nato pa se zmanjšuje. Posledice diferencialne rasti so značilne spremembe v obliki in sestavi telesa.
Alometrične enačbe
Hitrost rasti dela, komponente ali organa, y, lahko prikažemo relativno do drugih delov telesa ali celega telesa, x (preko celega obdobja rasti). Razmerje hitrosti rasti je propocionalno razmerju velikosti delov,
(dy/dx) = b(y/x)
To lahko zapišemo v enačbi, ki ima obliko y = axb. Ta enačba se imenuje alometrična enačba v kateri je b "rastni koeficient". Če je b=1, potem rasteta del y in x proporcionalno in ni spremembe v obliki. Če je b>1 ali če je b<1 potem del raste hitreje ali počasneje kot ostali in pride med rastjo do spremembe v obliki.
RASTNI CENTRI: AKTIVNI V RAZLIČNEM ČASU; RAZLIKE V AKTIVNOSTI; KOORDINIRANI; KARAKTERISTIKE VRSTE
MAKSIMALNA RAST: DOLOČEN VRSTNI RED, RAST DOLOČENEGA DELA ŽIVALI JE KONTROLIRANA Z AKTIVNOSTJO DRUGEGA DELA.
Relativna rastGenetsko standardizirana alometrična enačba
I = aTb,
kjer je I posamezen del, v T pa so upoštevani vsi deli.
Pri zrelosti,
Im= aTmb
a = Im/Tmb
I = (Im/Tmb) Tb
(I/Im) = (T/Tm)b
to je
uI= uT bEnačba uI= uT je genetsko standardizirana oblika alometrične enačbe. I in Im ter T in Tm imajo vedno enake enote zato je enačba brez enote, b (razmerje relativnih hitrosti rasti) pa je vedno stndardizirano in primerljivo za vse vrste in pasme. Grafično lahko alometrično enačbo prikažemo v alometričnem kvadratu.
PREDIKTIVNE ALOMETRIČNE ENAČBE ( X = klavna masa)KOSTI: Y1 = 0.077 A0.26 X0.74
MIŠIČJE: Y2 = 0.35 A0.02 X0.88
MAŠČOBNO TKIVO: Y3 = 0.19 A- 0.65 X1.65Relativna zrelost tkiv
Stopnja zrelosti mišic, maščobnega tkiva in kosti po disekciji v razmerju s stopnjo zrelosti celega telesa (Butterfield in sod., 1983 (Anim. Prod.)).
| Tel.deli | Stopnja zrelosti | ||||||||||
| Celo telo | 0 | 0.1 | 0.20 | 0.30 | 0.40 | 0.50 | 0.60 | 0.70 | 0.80 | 0.90 | 1.00 |
| Mišičevje | 0 | - | 0.24 | 0.35 | 0.46 | 0.56 | 0.66 | 0.75 | 0.84 | 0.92 | 1.00 |
| Mašč.tkivo | 0 | - | 0.05 | 0.11 | 0.18 | 0.27 | 0.38 | 0.51 | 0.65 | 0.82 | 1.00 |
| Kosti | 0 | - | 0.26 | 0.34 | 0.50 | 0.60 | 0.70 | 0.79 | 0.86 | 0.94 | 1.00 |
| . |
|
|
|||
| Vrsta |
|
|
|
|
|
| Tele |
|
|
|
|
|
| Mačka |
|
|
|
|
|
| Mor. prašiček |
|
|
|
|
|
| Jagnje |
|
|
|
|
|
| Človek |
|
|
|
|
|
| Miš |
|
|
|
|
|
| Kunec |
|
|
|
|
|
| Podgana |
|
|
|
|
|
| Prašič |
|
|
|
|
|
Presenetljive so razlike tako v rojstni masi kot v telesni sestavi ob rojstvu - posebno v količini maščobe. Tako ima prašič pri rojstvu le 1.1% maščobe, morski prašiček in človek pa okrog 10%.Spremembe kemične sestave telesa med rastjo tipičnega sesalca
Povprečna kemijska sestava (odraslega) telesa po Mitchellu (1962) je naslednja:
voda 58.6%
lipidi 18.4%
proteini 18.4%
pepel 4.6%.Kemijska telesna sestava dozori, ko žival v telesni masi doseže stopnjo zrelosti približno 0.3. Rahlo narašča le delež prepela.
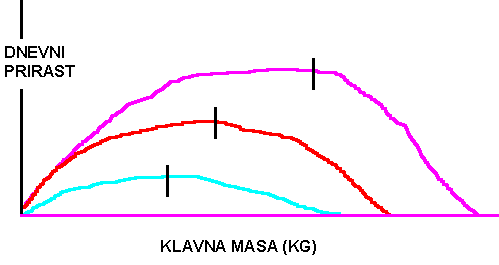
POTEK IN ZMOGLJIVOST RASTI (PRIMER PRAŠIČI) RASTNOST - prirast teže v časovni enoti. Zanimive so razlike v rastnosti med pasmami in med živalmi znotraj ene pasme.
POTEK RASTI - Pojem opredeljuje spreminjanje hitrosti rasti v posameznih obdobjih (npr. živali določene pasme rastejo v skladu z določeno rastno krivuljo; razlike v rastnih krivuljah med pasmami in znotraj pasem pa so značilne.
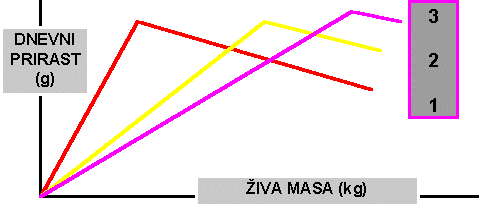
Potek rasti prašičev od 30 do 100kg, ki je pokazana v grafu, je zelo različna. Prašič številka ena je imel veliko rastnost na začetku rasti, pri večji teži pa je bila rast manjša. Taki prašiči so običajno zelo zamaščeni in porabijo za enoto prirasta veliko krme. Povsem drugače pa je potekala rast prašiča številka tri. Počasnejša rast na začetku in hitrejša rast pri večjih težah. Taki prašiči so običajno mesnatejši in v vsakem primeru porabijo manj krme za enoto prirasta.Trije poteki rasti prašičev
ZMOGLJIVOST RASTI pomeni končno velikost živali, pojem lahko uporabimo tudi za posamezna tkiva; zelo zanimiva je zmogljivost rasti mišičnega tkiva (velike razlike ne samo med pasmami, ampak tudi med živalmi znotraj ene pasme).
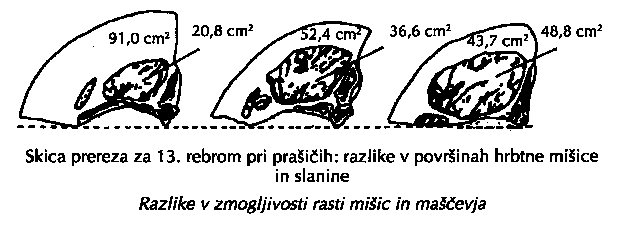
Prašiči iz gornje slike so bili ob zakolu enako težki (107 kg). Med njimi pa so izredno velike razlike v sestavi telesa. Ker so bili krmljeni enako intenzivno z enako krmo in so enake pasme ter spola, potem lahko zaključujemo, da so razlike v mesnatosti posledica različne zmogljivosti rasti posameznih telesnih tkiv. To pa je samo en dokaz več, kako pomembno je, da za pleme poiščemo in izberemo take živali, ki imajo veliko zmogljivost rasti mišičja.V literaturi je le nekaj informacij o selekciji poteka in zmogljivosti rasti živali. Eisen (1976) poroča, da so sicer maloštevilni poskusi pri miših pokazali srednjo uspešnost selekcije spremembe poteka rasti krivulje. Šalehar (1978, 1979) je v raziskavah rasti prašičev potrdil razlike v poteku rasti med pasmami in opredelil njegov pomen za selekcijo rasti prašičev. Krieter (1987) je na vsakoletnem simpoziju kmetijske fakultete v Kielu v predavanju "Zuchterische Moglichkeiten zur Regulation des Wachtstums beim Schwein" predstavil zanimive predhodne rezultate raziskave rasti prašičev nemška plemenita in Pietrain. Srednje vrednosti za parametre rastne krivulje in nekatere mere rasti pri obeh pasmah so v tabeli 3.
| . |
|
|
| končna masa (kg) |
|
|
| merilo zrelosti |
|
|
| masa pri max.prirastu (kg) |
|
|
| Starost pri maks. prirastu (teden) |
|
|
| Maks.prirast pri inflekciji (kg/teden) |
|
|
Končna masa je med pasmami različna. Pasma pietrain je primer rano zrele in bela plemenita pozno zrele pasme. To potrjujejo tudi podatki za mere rasti in mere rastne krivulje.Korelacije med parametri krivulje in dnevnim prirastom (Krieter, 1988)
A k y t v Dnevni prirast
A - -.82 .82 .76 -.02 -.20
k - -.47 -.57 .52 .68
y - .91 .36 .02
t - .08 -.19
v - .83
A - končna masa
k - merilo zrelosti
y - masa pri maks. prirastu
t - starost pri maks. prirastu
v - maks. prirast pri inflekcijiMed dnevnim prirastom in končno maso ter starostjo pri maksimalnem prirastu je negativna korelacija. Živali z veliko zmogljivostjo rasti - veliko končno maso, so težje in starejše pri maksimalnem prirastu. Visoki in pozitivni korelaciji sta med dnevnim prirastom in merilom zrelosti ter maksimalnem prirastu pri inflekciji. To omogoča sklepanje na splošno manjšo rastnost živali z veliko zmogljivostjo rasti.
Tabela : Korelacija med parametri krivulje in kriteriji selekcije (Krieter, 1987)
SESTAVA TELESA DOLOČEN VRSTNI RED RASTI TELESNIH TKIV:
ŽIVČEVJE --> KOSTI -->MIŠICE --> MAŠČOBNO TKIVO
POSLEDICE:
SPREMINJANJE SESTAVE PRIRASTA
SPREMINJANJE SESTAVE TELESAKEMIČNA SESTAVA TELESA KEMIČNA SESTAVA TELESA PRAKTIČNO DOSEŽE ZRELOST, KO JE STOPNJA ZRELOSTI TELESNE MASE CCA 0.3; DELEŽ PEPELA ŠE NEKOLIKO NARAŠČA. NA BREZ MAŠČOBNI OSNOVI JE KEMIČNA SESTAVA TELESA PRAKTIČNO KONSTANTNA:
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|